All published articles of this journal are available on ScienceDirect.

Effectiveness of Protected Areas in Conserving the Highly Hunted Mammal Species as Bushmeat in Southern Benin
Authors Info & Affiliations
Abstract
Background:
The habitat degradation together with fragmentation and illegal hunting represent a major threat for biodiversity conservation in Lama protected areas.
Method:
We used a combination of questionnaire survey with local communities for ranking the hunted mammal species as bushmeat and track surveys in gridded-cell system of 500x500 m2 (n=268) to assess at what extend the management design, the anthropogenic factors and habitat type affect the occupancy model of those mammal species.
Results:
Twenty mammal species have been predominantly reported by the local inhabitants to consume bushmeat species and 5 of them have been identified as the most preferable as hunted game mammals. The selection of the preferred habitat among the swampy forest, the dense forest, the tree plantations and cropland for the prioritized game species varies between species but looks similar when grouping in different orders. Some bushmeat species were found to select the more secure habitat (natural forest); suggesting the zoning system in the Lama forest can passively protect those species. However, some species such as T. swinderianus although highly hunted showed preference to anthropogenic habitat, avoiding the well secured core zone in Lama Forest.
Conclusion:
Our findings highlighted the importance of the zoning system with different management objectives in the habitat occupancy model of the highly hunted wildlife species.
1. INTRODUCTION
The sustainability of bushmeat species in Central and West Africa is well documented in the literature (Fa and Brown 2009). The poaching for trade is much more preferred in the remote forest habitat because of increase in penetration logging roads (Wilkie et al. 2000). This was exacerbated by the fact that poachers use the sophisticate greater access to modern weapons (Bowen-Jones and Pendry 1999). Additionally, poverty and lack of protein demand for food could explain the illegal hunting activities by the local populations living around the protected areas, they can consume the bushmeat either directly or sell for some incomes (Wilkie et al. 2005, Hema et al. 2017). In such way, the Protected Areas (PAs) could be viewed as an important reservoir in many regions (Fa et al. 2006).
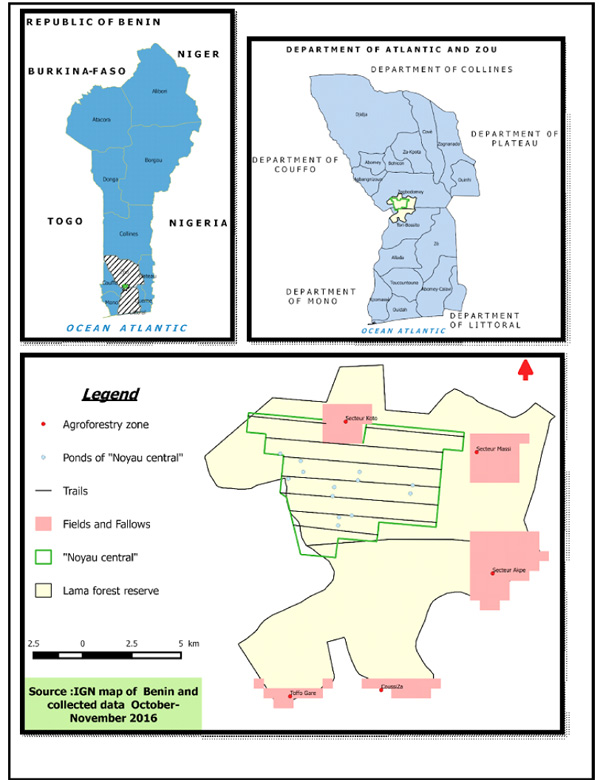
Globally, the PAs play a greater role in conservation strategies by helping to prevent at some extend from biodiversity loss (Bruner et al. 2001). But research on how far the PAs can help to protect wildlife from bushmeat poached and some other threats is limited (Gaston et al. 2008). According to the MAB-UNESCO design, the classical PA mostly has three components with different management objectives, those are: core zones, a buffer zone, and a transition zone. Till 2009, this model of zoning has been adopted by 170 countries worldwide totalizing 553 protected areas (Roe et al. 2009). and this number is still increasing every year (Roe et al. 2009). The Lama forest holds this same design management system with a buffer zone mainly harboring an important teak plantation, which is exploited by the National Office of Wood production (ONAB), and the core zone strictly protected by ONAB. This forest has been destroyed by the local farmers during the period 1946-1986, in search for more fertilized land and also due to the lack of a management plan of the forest at that period. Consequently, the original big area of the natural semi deciduous forest has been fragmented in Lama region into a small stand of natural semi deciduous forest remains in 1987 throughout the protected forest. After putting the management of the Lama protected forest in the charge of the ONAB authority, the destruction of all the huts inside the remnant forest was carried out systematically in January 1988. Regarding the zoning plan, the displacement of the local population from some illegally occupied zones allowed to recover up to 4,777 ha of natural semi deciduous forest named the core zone or “Noyau Central” which is strictly managed for scientific research and biodiversity conservation concerns. Rangers trained and employed by the ONAB to secure the “Noyau Central” implemented regular anti-poaching activities. The remaining degraded part of the forest was divided according to different uses: 7,000 ha for timber production, 2,500 ha for fuel wood production and to resettle the population who was evicted from the forest and design as a forest plantation zone. Evicted farmers were posted in the management fields now called agroforestry centers, designed as the buffer zone. Income generated from the plantation exploitation is somehow used to secure the core zone which is a natural semi deciduous forest, very important for the wildlife conservation and the regeneration of natural forest (Nagel et al. 2004). In view of the most endemic and endangered animal species, the core zone is the highly suitable habitat, and is strictly protected according to well defined conservation objectives and should be a typical example of natural or minimally disturbed ecosystems (Augustin et al.1996, WU Wang 2004).
Enforcement of hunting restrictions in the human settlements surrounding tropical forests is difficult, perhaps unrealistic, and even socially undesirable, as long as the current socioeconomic conditions persist (Carrillo et al. 2000, Licona et al. 2011). Yet overexploitation must be avoided and hunting should be sustainable to prevent from extinction of many large mammal in the Lama forest. . Indeed hunting for bushmeat has led to several species depletion and even local extinctions (Bassett 2005, Cowlishaw et al. 2005, Fa et al. 2006). Achieving this goal is possible only if we have some scientific based information about the effectiveness of the management design in protecting the wildlife species against the multitude of threats such as the bushmeat hunting that they are currently facing (Djagoun and Gaubert 2009, Petrozzi et al. 2016). To our knowledge, this study is a first comprehensive attempt to evaluate how different management designation, anthropogenic and habitat characteristics influenced the occurrence of wildlife species in tropical forest.
Nowadays, the participative management which meets the biodiversity preservation goals and local population needs is promoted by delineating multiple-use buffer zones surrounding the strictly protected core zones (Peres and Zimmerman 2001). However, coping with the protection objectives of wildlife and the sustainable use are challenging the protected area management in Lama forest. In such consideration (i) do animals always recognize the more secure zone and avoid the buffer zones and transition zones? (ii) Can we assume that this zoning model worldwide accepted in the PA management does show a benefit of passively protecting wildlife in the wildlife population?
Information related to the effectiveness of the protected areas management design on the conservation of the wildlife species could help to put further management recommendation of Lama forest in southern Benin where hunting of wild animals is an important component of household economies (Djagoun and Gaubert 2009). In such consideration, it is important to evaluate the current management efficacy efforts to well preserve the biodiversity. Furthermore, we need to determine and address the factors that are influencing the presence and distribution of animals in this ecosystem. The goals of this study are to 1) determine the prioritized hunting bushmeat species in Lama forest and check if the management designation influences the occupancy of highly hunted wildlife animal around Lama forest; 2) identify the habitat and the environmental characteristics with the greatest effects on the wildlife species; and 3) make recommendations for wildlife conservation at the interface of the PA management and bushmeat trade.
2. METHODS
2.1. Study Area
Field surveys were conducted in the Lama forest and its surrounding villages. Lama forest is located in the Sudano-Guinean climatic zone which characterizes southern Benin (White 1986) (Fig. 1). The specificity of the southern Benin area is its situation in the Dahomey Gap zone (Nagel et al. 2004). The forest benefit from a subequatorial climate with two unequal rainy seasons alternating with two dry seasons. The annual rainfall is 1100 mm with a maximum evaporation of 500 mm, and the annual average temperature is 29.9°C (ranging between 22.38°C and 31.58°C). The Lama forest (4777 ha) is subdivided into three main zones: the natural forest, which is strictly protected (the so called “Noyau central”), the forest plantations considered as the buffer zone, and the transition zone, where local communities are allowed to use natural resources and farm. Several emblematic species of mammals are encountered in this forest, including the mona monkey (Cercopithecus mona), the red-bellied monkey (Cercopithecus erythrogaster erythrogaster), which is endemic to the Dahomey Gap, the vervet monkey (Cercopithecus aethiops), and threatened ungulates, such as the sitatunga (Tragelaphus spekei), the royal antelope (Neotragus pygmaeus), the black duiker (Cephalophus niger), and the yellow-backed duiker (Cephalophus silvicultor) (Kassa 2001, Sinsin et al. 2002). The dominant ethnic groups living around the Lama forest are, respectively, the Holli and the Fon.
2.2. Bushmeat Hunter Surveys
Only the cooperating hunters were interviewed to collect hunting information and this in six villages (Tègon, Koussi, Sèhouè, Massi, Koto and Hlagbadénou-Hlagbalonmè) surrounding the Lama forest from January to March 2015. We were accompanied by a local assistant who can speak the local language (holli or fon) to facilitate our contact with the hunters and collect more reliable data, in combining fixed-response and open-ended questions. Questions were related to the household size, number of owned domestic animals, hunting methods, total number of animals hunted per species. A total of 80 active hunters participated to questionnaire and 65 hunters responded to all questionnaires and were consistently interviewed every week during the field work period for recording the amount of catch.
2.3. Habitat Occupancy Surveys
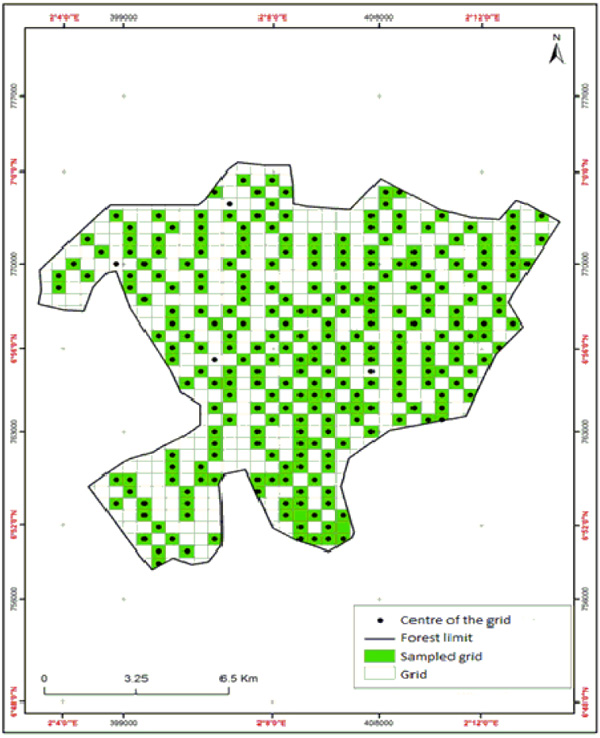
An occupancy survey was conducted from December 2014 to March 2015 to assess the occurrence of the most hunting bushmeat species in the Lama forest. A grid-cell of 250,000 m2 was used as the sampling unit (Fig. 2). A total number of 673 grids were generated covering different habitat types and different management zone in Lama forest using a GIS software. We labeled each grid cell using a unique ID whereas 268 grids were selected randomly, and their respective GPS coordinates noted. We assume that animal movement from site to site within the Lama forest is on random basis and if so the computing the proportion of the sampled grid cell used by the animals as occupancy estimator remained unbiased (MacKenzie and Royle 2005). Additionally, we assumed that all sampling grid to be in detection probability, heterogeneous (Royle and Nichols 2003). Three survey teams were established to help in data collection which included three persons per team. The team leader was a university-trained person team and two local assistants selected from 10 years skilled local hunters. A short training was given to the teams regarding the survey techniques and most essentially how to identify the Lama forest's animal tracks and dung based on shape and size. We took one day as test to complete the grid survey together to make some adjustment regarding the animal signs identification, precisely filling of data forms, and use of the GPS in go to function to find the selected sampling grids. After this test, the teams were launched to investigate each selected sampling grid, searching for presence of the targeted bushmeat species, according to the observation of dung and tracks fresh. But also when possible the direct observations of animals were recorded. For subsequent analysis, coding “0” for absence and “1” for presence generated presence/absence data of the species in each grid. A plot of 30 m X 30 m was set up in the middle of each sampling grids to record habitat (“Dense forest”, “Swampy forest”, “Cropland”, “Forest plantation”) and environment characteristic. The recorded variables were: tree dbh (A), canopy cover (B), zone (C), nearest distance to village (D), nearest distance to road (E), under canopy cover (F) nearest distance to water point (G), number of trees (H).
2.4. Data Analysis
Hunting data analysis was partly based on Martin (1995) method which supports that when people are asked to freely recall things, they tend to list the most significant one first (Assogbadjo et al. 2012). In addition, almost everybody cites prominent categories, while a minority of hunters mentions less significant ones. The most hunted species ranked by hunters were assessed in Lama forest by computing the average order in which each specie was cited by adding together their respective rank according to the hunters and finally this was divided by the total number of hunters participating to the survey (65 individuals in total). This value could be greater for given species when a couple of respondents have cited the bushmeat species as important. Results from this analysis are displayed on figures for visualization.
Bushmeat species habitat preference indices (Manly’s alpha) for the four different habitat types encountered for Lama forest were calculated by using the Manly et al. (1972) approaches and the formula is shown below:
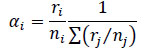 |
where
αi Manly’s α value for habitat i
ri, rj proportion of habitat type i or j used (i and j=1, 2, 3, …, m)
ni, nj proportion of habitat type i or j available in the study area, m represents the number of total habitat type available in the study area (m=4 in this study). The proportion of habitat type i or j availability was performed in making the ratio between total surface of habitat type i or j available and the total surface of the Lama forest using the shape file of habitat type in lama forest with ArcGIS software. While the proportion of habitat type i or j used represents the number of time a given animal has been observed in habitat type i or j divided by overall number of observation. When there is no preference in habitat selection αi = m–1. If αi> m–1, habitat type i is preferred while αi < m–1 indicates avoidance of habitat type i. This analysis allows investigating change in habitat selection across protected zone and non-protected zones (“Dense forest”, “Swampy forest”, “Cropland”, “Forest plantation”).
We estimated resource-selection functions (RSF’s) (Manly et al. 2002) following the Generalized Linear Model with logistic family to identify which zone (Natural forest, the forest plantation, and the buffer zone) and environmental factors were associated with the occurrence of bushmeat species. Estimation of parameters was carried out using the function glm in the R software package. The procedure of backward stepwise was used for modelling simplification and the model was selected with the lowest AIC (Akaike Information Criterion) values. The predictors are: tree dbh (A), canopy cover (B), zone (C: Natural forest, the forest plantation, and the buffer zone), nearest distance to village (D), nearest distance to road (E), under canopy cover (F) nearest distance to water point (G), number of trees (H) and the response variable is the presence/absence data of the species in each grid were generated by coding 0 for absence and 1 for presence. Models with ΔAIC scores within 2 units from the most parsimonious model were included as alternative models. The Pearson correlation coefficient was used to examine the co-linearity among the predictor variables. All analyses were performed in the statistical program R version 2.14.0. (R Development Core Team 2012).
3. RESULTS
3.1. The Mostly Hunted Bushmeat Species in Lama Forest
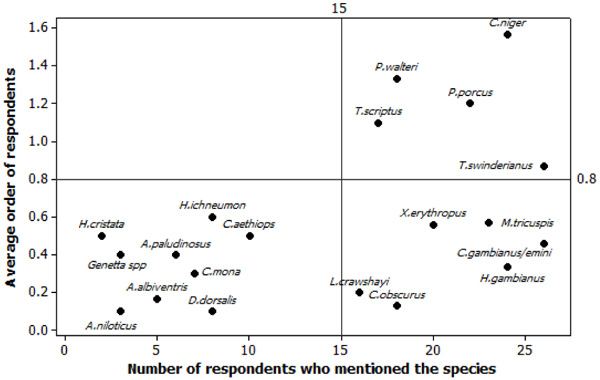
We have recorded twenty species as the most preferred bushmeat species. The recorded species can be grouped into three categories. The first category encompasses the mostly cited wildlife as bushmeat species with high hunting pressure (Fig. 3). Among this category, we recorded ungulates such as the black duiker (Cephalophus niger), the blue duiker (Philantomba walteri) (Fig. 4a), the Red river hog (Potamochoerus porcus) and bushbuck (Tragelaphus scriptus) (Fig. 4c). The cane rat (Thryonomys swinderianus) (Fig. 4b), also belongs to this category despite it is less preferred than the previous species cited. The Gambian rat (Cricetomys gambianus and C. emini), the cusimanse (Chrossarchus obscurus), the pangolin (Manis tricuspis) and the squirrel (Xerus erythropus) (Fig. 4d) were recorded in the second category: the species highly mentioned by the hunters but with lower rank according to the priority in hunting. The third category of bushmeat species is the one less cited by the respondents and not receiving priority in terms of hunting. This class included mostly the small carnivores species (Genetta spp, Herpestes ichneumon, Atilax paludinosus) and some primate species (Cercopithecus aethiops, Cercopithecus mona).
3.2. Habitat Use Assessment of the Most Hunted Bushmeat Species
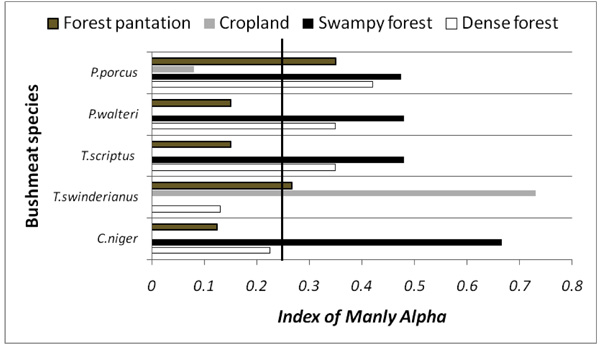
Fig. (5) reports the manly alpha index calculated for the most hunted species in Lama Forest. The rodent species (T. swinderianus) mostly selected the tree plantation and the cropland in Lama forest (αi > 0.25), but avoid the other habitats. However, the bushbuck (T. scriptus), black duiker (C. niger) and blue duiker (P. walteri) preferred the dense and swampy forests while avoiding the plantation and cropland. The red river hog (P. porcus), mostly used the dense and the swampy forests.
3.3. Modeling the Mostly Hunted Bushmeat Species in Lama Forest
The most parsimonious candidate models for each species are presented for each species with the ΔAIC indicating the ‘best-model’ (i.e. lowest AIC value) and the alternative models at ΔAIC˂ 2 (Table 1). The details are given in Table 2 regarding the best model predicting habitat occurrence per species. Our results show that the occurrence area of T. scriptus can be well predicted by the distance to the water point, the canopy cover with a significant preference to the core zone in Lama forest (β=0.62; p=0.02). A good condition of canopy cover (β=3.05; p<0.001) and a close distance to the village (β=0.65; p=0.01) well predict the occurrence of T. swinderianus in the Lama forest. The tree diameter significantly predicts the presence of C. niger with less presence at proximity of the trees having large diameter (β=-0.09; p<0.001). P. walteri significantly occurs in the protected zone (β=-4.32; p=0.02). In the final model retained for P. porcus, a significant positive selection was found for distance to water point (β=3.12; p=0.03) and the core zone (β=5.32; <0.001).
| Models | AICc | ΔAICc |
|---|---|---|
| 1- T.scriptus | ||
| p(ABGC) | 77.95 | 0 |
| p(GC) | 78.91 | 0.96 |
| p(HFC) | 79.56 | 1.61 |
| 2- T. swinderianus | ||
| p(DCF) | 51.63 | 0 |
| p(ABGC) | 53.12 | 1.49 |
| 3- C. niger | ||
| p(ABG) | 17.56 | 0 |
| p(GC) | 18.87 | 1.31 |
| 4- P.walteri | ||
| p(HFC) | 10.43 | 0 |
| p(DCF) | 12.03 | 1.6 |
| p(FC) | 12.31 | 1.88 |
| 5- P.porcus | ||
| p(GCD) | 46.35 | 0 |
| Species | Variables | Estimate(β) | Std. Error | P-value | aAIC |
|---|---|---|---|---|---|
| T.scriptus | intercept | 2.32 | 2.03 | 0.36 | - |
| - | dbh | -0.57 | 1.68 | 0.09 | - |
| - | canopy | 1.43 | 0.97 | <0.001 | 77.95 |
| - | natural.forest | 0.62 | 0.35 | 0.02 | - |
| - | dist.water | -0.22 | 1.35 | 0.01 | - |
| T. swinderianus | intercept | -5.15 | 3.56 | 0.09 | - |
| - | canopy | 3.05 | 2.56 | <0.001 | 51.63 |
| - | natural.forest | -0.92 | 0.66 | 0.06 | - |
| - | dist.village | 0.65 | 0.46 | 0.01 | - |
| C.niger | intercept | 1.03 | 2.15 | 0.07 | - |
| - | dbh | -0.09 | 0.05 | <0.001 | 17.56 |
| - | canopy | 0.82 | 1.20 | 0.75 | - |
| - | dist.water | 0.07 | 1.05 | 0.45 | - |
| P.walteri | intercept | 3.12 | 3.04 | 0.04 | - |
| - | forest.plantation | 4.32 | 4.02 | 0.02 | 10.43 |
| - | under.canopy | -0.97 | 1.96 | 0.96 | - |
| - | nbr.trees | 1.36 | 1.06 | 0.06 | - |
| P.porcus | intercept | -7.38 | 6.38 | 0.89 | |
| - | natural.forest | 5.32 | 4.68 | <0.001 | 46.35 |
| - | dist.village | -2.13 | 1.98 | 0.06 | - |
| - | dist.water | 3.12 | 3.08 | 0.03 | - |
| aThe lowest AIC value indicating the best model as reported in Table (1) | |||||
4. DISCUSSION
The importance of mammal populations in providing an important source of cash income and protein to local communities is well documented worldwide (Fitzgibbon et al. 1995). The bushmeat trade is dominated by ungulates and rodents in southern Benin like in several other places in West and Central Africa (Bassett 2005, Fa and Brown 2009).
Hunting restrictions enforcement within local population surrounding the tropical forests is difficult, and may be unrealistic regarding the persisting very worse socioeconomic conditions (Licona et al. 2011). Yet overexploitation must be avoided and hunting should be sustainable to prevent from the population decreasing of many large animals or drastically their extinction in the Lama forest. Indeed hunting for bushmeat has led to several species depletion and even local extinctions (Bassett 2005, Cowlishaw et al. 2005, Fa et al. 2006, Akani et al. 2015). Wildlife abundance and richness constitute one of the factors explaining the hunting of the wildlife for trading around the protected areas. Although the better conditions offered by a given strictly protected area it is however possible to have the buffer zone and the areas outside the protected area as richest sources of protein. Our findings on the mostly hunted bushmeat species in Lama forest showed that some of the species observed in the bushmeat markets such as C. niger, P. walteri, P. porcus and T. scriptus are known to occur almost exclusively within the Lama forest, especially in the core zone which is strictly protected (Kassa 2001). We also checked the habitat selection of those commonly hunted species to see if they select habitats outside Lama Forest despite the security offered in the core zone. We recorded less occurrence data for the species traditionally preferred by hunters in the buffer zone. The four species listed above were found to select their habitat according to the vegetation cover, trees density, distance to the water point and the limited human impact. Only the natural forest located in the core zone can offer such conditions and this confirms our results of less occurrence outside the core zone.
Similarly, core protected zone within the Biosphere Reserve design has also been reported in Brazil and Cameroon to harbor more higher ungulate population density compared with more human accessible outer areas (Peres and Zimmerman 2001). Bruner et al. (2001) revealed the same finding as ours when reporting that most of the 93 tropical protected areas they studied are facing smaller decreasing in game populations than the surrounding areas. However, the penetration in protected areas themselves for hunting and the emptying of areas surrounding protected areas suggest that the wildlife populations are not sustainably preserved in protected areas.
The cane rat (T. swinderianus) also highly hunted in the Lama forest, showed preference for the buffer zone and areas close to the village. However, rodents are in fact reported to forage mostly in the in anthropogenic disturbed areas (Eves and Ruggiero 2000). Degraded secondary vegetation, particularly when interspersed with agricultural fields, may provide adequate habitat for rodent species (Muchaal and Ngandjui 1999) such as T. swinderianus and could account for its apparently similar distribution across the study site regardless of hunting pressures.
The management model of the Lama forest is similar to the one of Biosphere Reserve with a core zone with is the forest, a buffer zone which is the tree plantation and the villages. The benefits of passive wildlife protection according to the Biosphere Reserve model adopted in Lama Forest management system. Despite the participative management approaches, the management design of Biosphere Reserve has been championed in including indigenous peoples and their ecological knowledge in co-managing the surrounding resources (Berkes 2004), an evidence of illegal hunting activities has been recorded in the in the core zone of Lama forest, although strictly protected. In case of passive management the buffering concept of the Biosphere Reserve model, these approaches showed their limit and the necessity to reinforce anti-poaching strategies in the protected areas is strongly needed. According to Bruner et al. (2001), the needs to clearly agreed with all conservation stakeholders, upon conservation goals as well as demarcated boundaries, public awareness of laws and the presence of guards appear to be very important in wildlife conservation. Implementing some economical generating income such as beep keeping, game farming could help to minimize their impact on the natural resources.
Interestingly, our study only provided limited information on how the basic habitat features of the Lama forest (tree dbh, canopy cover, management zone, nearest distance to village, nearest distance to road, under canopy cover, nearest distance to water point and number of trees) influence the occurrence of bushmeat species in Lama Forest. Many of habitat features assessed in this study showed some influence to the habitat occurrence of the bushmeat species, but this habitat features could fail in predicting well occurrence of the animal when considering the high anthropogenic factors occurring the Lama forest. Possibly, fine-scale measures of vegetative structure or microhabitat conditions not included in this study would have better elucidated the influences of habitat on wildlife occurrence in the Lama forest. Importantly, the occupancy modeling use here does not include the animal density difference within the sampling and recommended further model to take into account this difference for more accuracy in estimating the wildlife population density responses to the same environment variables measured in this study. However, the relatively low densities at which these the study animals occur in the Lama forest does not limit our findings of the predictors that affect the distribution of wildlife.
CONCLUSION
Overall, the results of the present study suggest that many mammal species are suggested to hunting around the Lama forest but also the importance of the zoning system with different management objectives in the habitat occupancy model of the highly hunted wildlife species was highlighted. Thus, careful field surveys remain essential in order to establish the presence/absence, and obviously also the relative abundance, of a considerable portion of the mammal fauna in the Lama forest to better show the benefits of passively protecting wildlife in the Lama forest.
CONSENT FOR PUBLICATION
Not applicable.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGMENTS
IDEA WILD supported the fieldwork equipment through the grant provided to Dr. Chabi A. M. Djagoun. Thank to Alexander von Humboldt Foundation through Georg Forster Research Fellowship (HERMES) provided to the first author to complete this manuscript writing. We are grateful to the local communities who participated in this research. Our acknowledgments also go to our field guide Mr. Lamidi, who has helped us for the field works. We also thank two anonymous reviewers for helpful comments on an earlier version of this manuscript.
REFERENCES
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link] [PubMed Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]
[CrossRef Link]

